Эмболомеры
Главные особенности
строения.
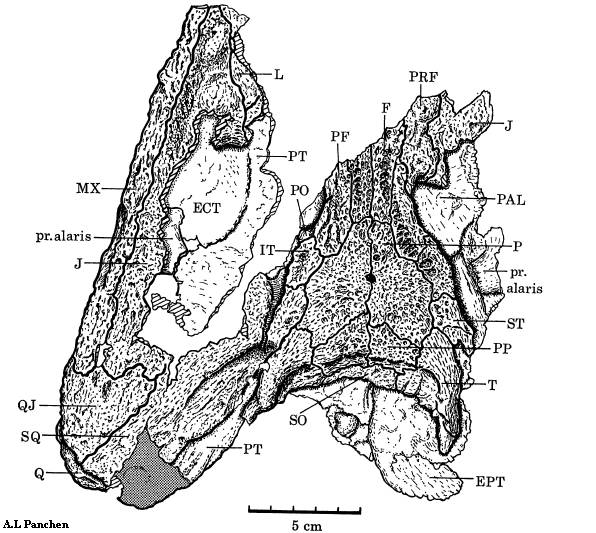
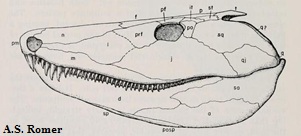
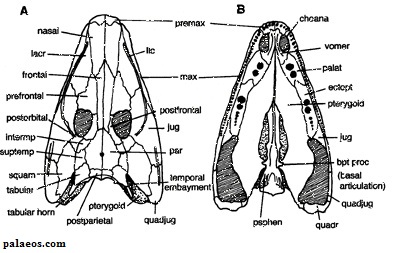
Рисунок 1. Череп Eoherpeton
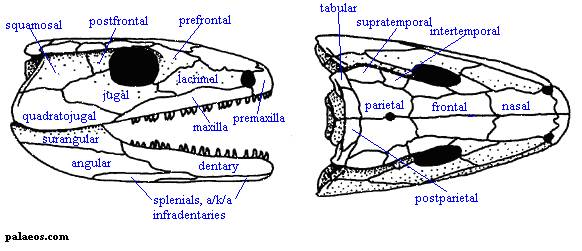
Термины: squamosal (чешуйчатая кость); postfrontal (заднелобная кость); supratemporal (надвисочная кость); parietal (теменная кость); quadratojugal (квадратноскуловая кость); tabular (таблитчатая кость); premaxilla (предчелюстная кость); maxilla (верхнечелюстная кость); lacrimal (слёзная кость); nasal (носовая кость); dentary (зубная кость); kinetic line (кинетическая линия, показана чёрным)
Основная характеристика черепа эмболомеров заключалась в
том, что его щёчная область (чешуйчатая и заднелобная кости) неплотно
соединялась с надвисочной костью. Линия между щёчной областью и остальным
черепом носит название кинетической; помимо эмболомеров, она также имелась у
кистепёрых рыб, предков амфибий.
_________________________________________________________________________
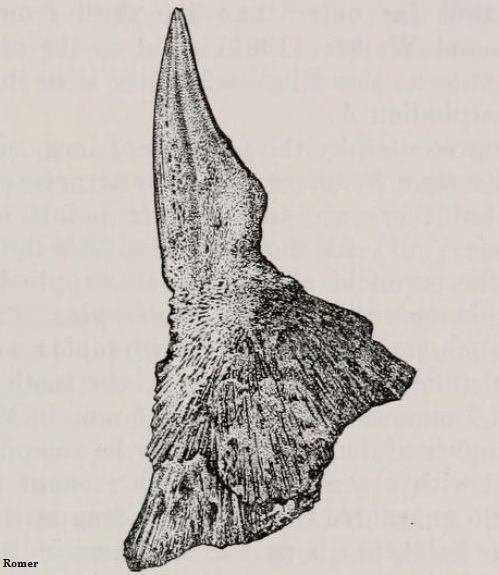
Рисунок 2. Зуб Anthracosaurus в разрезе
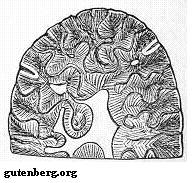
Эмболомеры имели лабиринтодонтные зубы, также как и темноспондилы, ихтиостегалии и кистепёрые рыбы. Внутри их зубов видна дентиновая складчатость.
_________________________________________________________________________
Рисунок 3. Передняя
конечность Proterogyrinus
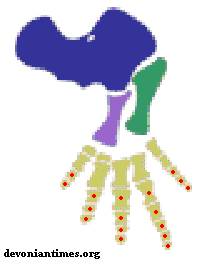
Термины: humerus (плечевая кость, показана синим цветом); radius (лучевая кость, показана сиреневым цветом); ulna (локтевая кость, показана зелёным цветом); фаланги пальцев отмечены красными точками.
Число пальцев эмболомеров - по 5 на передних и на задних конечностях. Фаланговая формула кисти обычно 2-3-4-5-3, но были и исключения.
_________________________________________________________________________
Рисунок 4. Позвонок Proterogyrinus
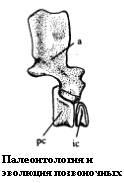
Термины: pc - pleurocentrum (плевроцентр); ic - intercentrum (гипоцентр); a - arcus neuralis (невральная дуга).
Эмболомеры имели примитивные для подкласса рептилиоморфов эмболомерные позвонки. В них с одной стороны располагался цилиндрический гипоцентр, а с другой - единый плевроцентр, который у первых амфибий и кистепёрых рыб был парным. У более развитых рептилиоморфов плевроцентр вытеснит гипоцентр, а у эмболомеров они занимают практически "равноправное" положение. Общее число предкрестцовых (то есть шейных и туловищных) позвонков эмболомеров было увеличено по сравнению с большинством других древних амфибий - примерно 40.
_________________________________________________________________________
_________________________________________________________________________
ГРУППЫ ЭМБОЛОМЕРОВ
Геохронологическая шкала:
http://evolbiol.ru/evidence05_files/eviden1.jpg
{kind=link}
Eoherpetontidae: Eoherpeton: E. watsoni PANCHEN
1975
Рисунок 5. Череп Eoherpeton - вид сзади
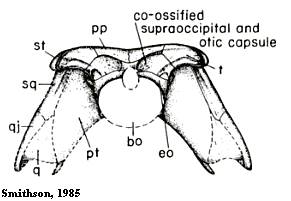
История изучения
Найденный череп, как предполагалось изначально (Watson 1929), принадлежал роду Pholidogaster (фолидогастер), отнесённому ныне к семейству колостеид, которые не принадлежали к эмболомерам и вообще рептилиоморфам. Позже окаменелости были переквалифицированы (Panchen 1975) и новый род получил название Eoherpeton (эогерпетон, "ранний гад"). Подробно он рассматривался также в работе другого автора (Smithson 1985).
Основные особенности
Раннекаменноугольный (визей - намюр) эмболомер из Великобритании - самый ранний из последевонских представителей отряда, опередивший более известный род Proterogyrinus на 8 миллионов лет. Единственный род и вид в своём семействе, полуназемное существо. Найден череп, а также позвонки и части поясов конечностей. Череп несколько удлинённой формы, с относительно круглыми орбитами и крупными ноздрями. Верхнечелюстная кость контактировала с квадратноскуловой (см. рисунок 1), это важная особенность эогерпетона. Другой его признак - отсутствие на черепе ушных вырезок. Что это такое, видно на рисунке 6.
Рисунок 6. Череп Palaeogyrinus
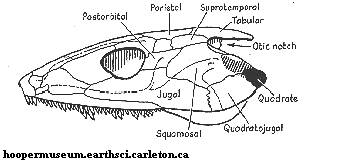
Термины: otic notch (ушная вырезка)
Длина черепа эогерпетона - 15 сантиметов, общая длина тела животного - примерно 1 метр. Имел цельный сильный таз, как у девонских амфибий (лобковая и седалищная кости крепко соединены друг с другом), и не в полной мере эмболомерные позвонки.
_________________________________________________________________________
Proterogyrinidae: Tulerpeton: T. curtum LEBEDEV
1984
Рисунок 7. Ископаемые останки Tulerpeton

Рисунок 8. Плечевой пояс Tulerpeton
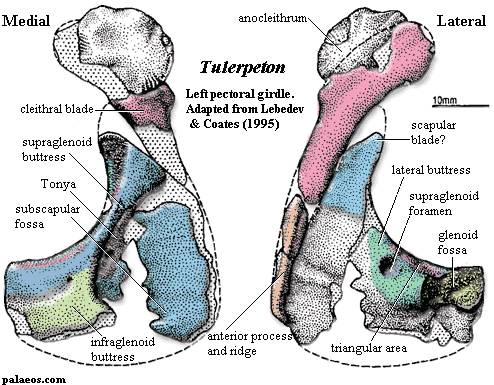
Термины: anocleithrum (аноклейтрум)
Рисунок 9. Конечности Tulerpeton
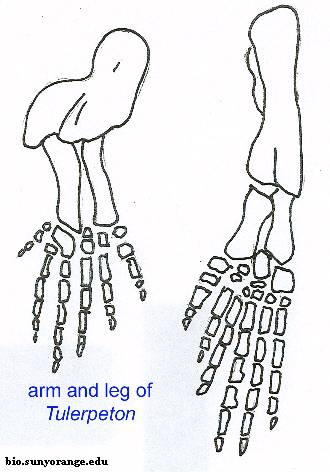
Рисунок 10. Плечевой
пояс Ichthyostega
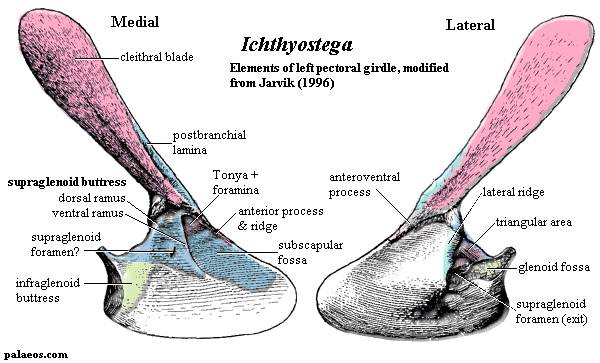
Термины: postbranchial lamina (медиальная пластинка клейтрума)
История изучения,
общие факты
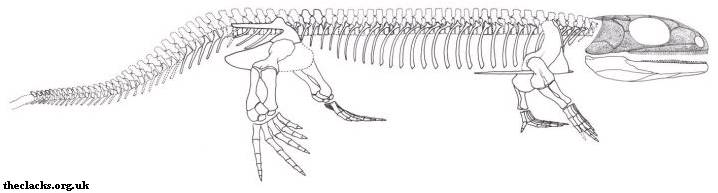
Tulerpeton (тулерпетон) обнаружен на территории Тульской области в России, отсюда и родовое название ("тульский гад"). Описан в работе Лебедева (Lebedev 1984), позже изучался другими ведущими специалистами по переходным формам от рыб к амфибиям. Наибольший интерес тулерпетон представляет как единственный эмболомер и вообще рептилиоморф девонского периода. Помимо всего прочего, окаменелости этого животного найдены в морских отложениях, а не в пресноводных, как у подавляющего большинства древних амфибий.
Основные особенности
Маленькое животное (около 60 сантиметров длиной). От черепа известны небольшие фрагменты (сошник и предчелюстная кость - см. рисунок 7), зато нормально сохранился посткраниальный скелет. Тулерпетона можно считать одной из наиболее изученных девонских амфибий, наряду с Ichthyostega (ихтиостега) и Acanthostega (акантостега). Не считая малоизвестную Jakubsonia (якубсония), тулерпетон является единственным получившим название девонским земноводным из России. Это также единственная амфибия, имевшая по шесть пальцев на передних конечностях и по семь на задних (ранее неверно считалось, что и на задних конечностях было по шесть пальцев; это отображено на рисунке 9). Данная черта отличает тулерпетона от всех остальных амфибий: у них было либо больше 6 пальцев на каждой конечности (ихтиостега, акантостега), либо 5 и меньше (Pederpes, Crassigyrinus и др.). Пальцы тулерпетона относительно тонкие и длинные, особенно на задних конечностях. В общих чертах строение плечевого пояса подобно рептиоморфному, за исключением одной детали: он сохранял аноклейтрум (см. рисунок 8) - очень примитивную кость, имевшуюся у акантостеги, но утраченную у ихтиостеги (см. рисунок 10). Как ни странно, аноклейтрум в виде исключения сохранялся и у некоторых других эмболомеров (Pholiderpeton). Так называемая медиальная пластинка клейтрума, сохранявшая у ихтиостеги (см. рисунок 10), у тулерпетона утрачена: это значит, у него не было внутренних жабр. Известны многочисленные рыбьи чешуйки на брюхе тулерпетона, защищавшие животное при передвижении. Несмотря на изученность вида, его систематическое положение точно не ясно. Тулерпетона иногда относят к рептилиоморфам, основываясь на тип строения его плечевого пояса. Некоторые учёные не согласны с такой классификацией; они считают тулерпетона более развитым, нежели ихтиостега и акантостега, но примитивнее всех остальных тетрапод. В качестве признака "crown group Tetrapoda", откуда исключают тулерпетона, указывается чаще всего наличие 5 пальцев.
_________________________________________________________________________
Proterogyrinidae: Papposaurus: P. traquairi
WATSON 1914
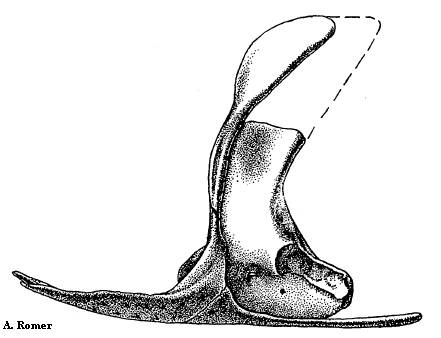
Рисунок 11. Бедро Papposaurus
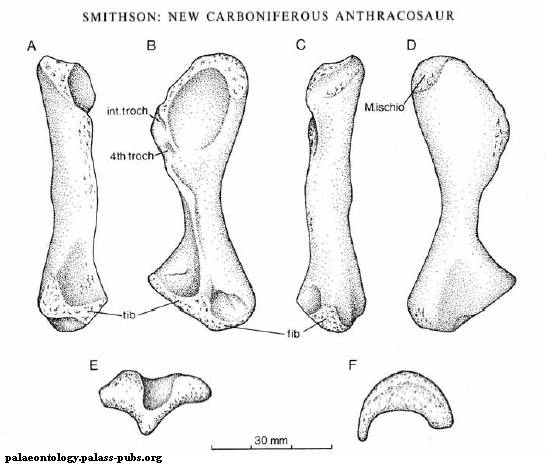
Возможно, Papposaurus traquairi - "пустое имя" (nomen vanum). Не исключено также, что это синоним более известного вида Proterogyrinus (протерогирин). Видовое название получил по фамилии учёного Р. Траквайра. Единственная окаменелость, приписываемая Papposaurus - правая бедренная кость из Шотландии, относящаяся к раннему карбону (миссисипию). В описании (Watson 1914) указывалось на сходство кости с таковой у ранних рептилий. Позже другими авторами (White 1939; Holmes 1984) у Papposaurus были найдены признаки, роднящие его с эмболомерами Archeria (арчерия) и Proterogyrinus. Бедро хорошо сохранилось, его длина - почти 7 сантиметров, оно было послабее, чем у арчерии.
_________________________________________________________________________
Proterogyrinidae: Proterogyrinus: P. scheelei
ROMER 1970 и P. pancheni SMITHSON 1986
Рисунок 12. Крыша черепа Proterogyrinus
pancheni
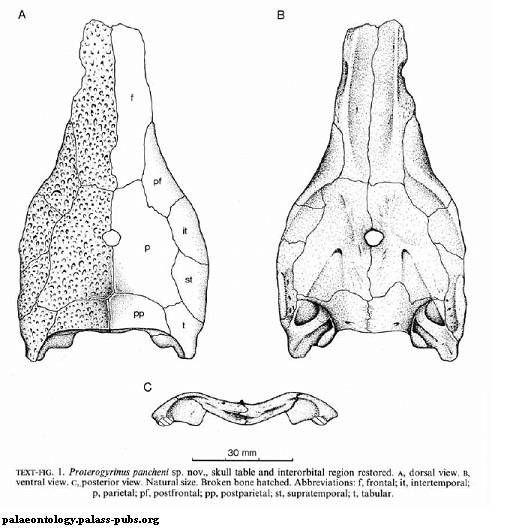
История изучения
Род Proterogyrinus
и типовой вид P. scheelei были названы А.
Ромером (Romer 1970). В
том же году был описан род Mauchchunkia
(Hotton 1970). Позже эти
названия были признаны синонимичными, и старшим синонимом оказался Proterogyrinus. P. scheelei также
рассматривался в работах Холмса (Holmes 1980 и 1984). Другой
вид протерогирина, P. pancheni (Smithson 1986), был назван в честь палеонтолога А. Панчена, который изучал
каменноугольных амфибий.
Общие факты
Один из самых примитивных и, в то же время, изученных эмболомеров. Несколько образцов типового вида найдены в Западной Виргинии (США), кости второго (нижняя челюсть, ребро и туловищный позвонок) - в Шотландии, как у множества эмболомеров. Протерогирин обитал на территории этих областей во время позднего карбона - серпуховского века. Название животного переводится как "ранний головастик". Вообще основы -gyrinus и -erpeton (от греч. ἑρπετόν - герпетон - "гад") чаще всего встречаются в названиях рептилиоморфов, как у рептилий -suchus и -saurus.
Основные особенности
Амфибия среднего размера (P. scheelei длиной до 1,5 метров, P. pancheni - до 1 метра). Длина черепа 12 - 20 сантиметров. Как и у большого количества вымерших земноводных, у типового вида протерогирина он имел каналы боковой линии. Что это такое, видно на снимке черепа Parotosuchus (не эмболомер) проекта Tree of Life:
Рисунок 13. Череп Parotosuchus

Протерогирин мог иметь барабанную перепонку, об этом говорит выемка на заднем крае черепа. Также найдено большое пластиновидное стремя (слуховая кость). По форме оно сильно отличается от тонкой слуховой косточки других животных. Ещё одна примитивная особенность черепа протерогирина - закрытое нёбо. Это видно на рисунке 14:
Рисунок 14. Нёбо Proterogyrinus
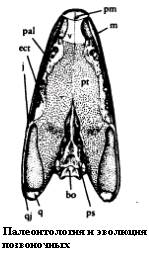
Характерной особенностью многих эмболомеров и протерогирина в их числе являлись два больших окна во внутренней стороне нижней челюсти. Их функция была, вероятно, связана с челюстной мускулатурой.
Рисунок 15. Нижняя
челюсть Proterogyrinus pancheni
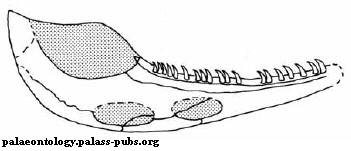
Термины: Meckelian fenestrae (на рисунке овальной формы, показаны серым цветом)
Отличия между видами протерогирина заключаются в строении зубов и позвоночника. Гипоцентры P. sheelei имели примитивную серповидную форму; плевроцентры были крупнее, именно они поддерживали невральную дугу (см. рисунок 4). Всего предкрестцовых позвонков у протерогирина 32 - меньше, чем у других эмболомеров. Конечности (схематично на рисунке 3) были относительно сильные; последние два факта говорят о более сухопутном образе жизни протерогирина, чем у его древнего родственника тулерпетона. Тем не менее, хвост протерогирина был длинный и уплощённый по краям, явно служивший для передвижения в воде.
______________________________________________________________________________________________
Proterogyrinidae: Silvanerpeton: S. miripedes
CLACK 1994
Рисунок 16. Скелет Silvanerpeton
Термины: interclavicle (межключица)
История изучения, общие
факты
Silvanerpeton (сильванерпетон) назван и описан Дж.А. Клак (Clack 1994) - специалистом по ранним тетраподам. Несколько лет назад этот вид также был рассмотрен в совместной работе Клак и Руты (Ruta & Clack 2006). Систематическое положение сильванерпетона точно не определено: возможно, это не протерогиринид, а базальный эмболомер, или даже примитивнейший из амниот. К последним относятся потомки рептилиоморфов - рептилии, птицы и млекопитающие. У всех них яйцо имеет амниотический мешок - полость, заполненную жидкостью, защищающей зародыш. Первые амниоты (Westlothiana и Casineria) жили уже в визейском веке, как и сильванерпетон.
Родовое название животного переводится как "лесной гад" (от Silvanus - римского бога леса). Животное с лесом никак не связано, а название такое получило из-за того, что человек, которые нашёл его окаменелости, имел фамилию Wood (англ. - "лес, древесина"). Игра слов иногда встречается и в названиях других родов. Так, например, родовое имя примитивного турнейского тетрапода Pederpes (кстати, тоже названного Дж. Клак) можно перевести и как "каменный гад", и как "стопа Педера". Видовое название S. miripedes переводится как "удивительная стопа".
Основные особенности
Несколько образцов составляют почти полный скелет. Более наземное существо, нежели протерогирин. Длина черепа - около 4,5 сантиметров, всего тела от рыла до таза - 20 сантиметров; длина хвоста точно неизвестна. Пятый палец стопы содержит дополнительную четвёртую фалангу, которой нет у родственников сильванерпетона, отсюда и название вида (см. выше). Фаланговая формула, таким образом, 2-3-4-5-4. Брюхо покрывает броня из взаимосвязанных чешуек. Похожая ситуация наблюдалась у многих амфибий, например, у девонского протерогиринида тулерпетона и у темноспондила Balanerpeton из той же области, что и сильванерпетон. Разве что у сильванерпетона эти чешуйки были потоньше. У него имелась также длинная заострённая межключица (см. рисунок 16), одна из костей плечевого пояса. Рёбра весьма удлинённые. Все известные кости сильванерпетона в таком виде, как они найдены, показаны на рисунке 17:
Рисунок 17. Кости Silvanerpeton
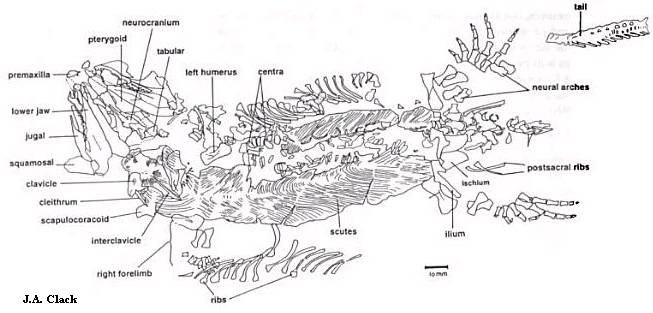
______________________________________________________________________________________________
Archeriidae: Archeria: A. crassidiscus CASE
1915
Рисунок 18. Череп Archeria.
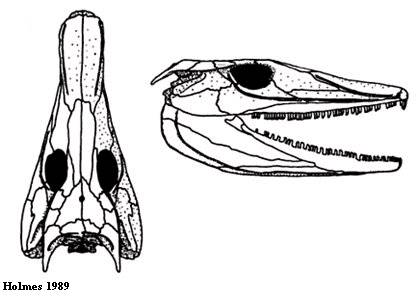
Рисунок 19. Череп Archeria.

Рисунок 20. Кисть Archeria
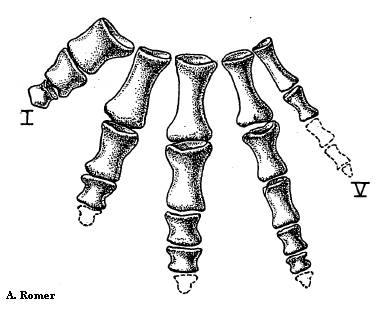
Рисунок 21. Плечевой
пояс Archeria
Термины: cleithrum (клейтрум, наверху)
Рисунок 22. Тела позвонков Archeria

История изучения
Достаточно известный эмболомер. Здесь рассмотрены только вопросы систематики вида. В 1884 году знаменитый палеонтолог Э. Коп определил разрозненные позвонки из ранней перми Техаса принадлежащими двум видам рода Cricotus (Сope 1884) - C. crassidiscus и C. hypantricus. Надо сказать, что сам род и некоторые его виды (в частности, C. heteroclitus) были названы Копом ещё в 1875, сейчас их систематическое положение не ясно. Позже (Case 1915) по нескольким хорошо сохранившимся пермским образцам, включая один почти полный скелет, был назван новый род Archeria (арчерия - в честь графства Арчер), включавший виды A. crassidiscus, A. robinsoni и A. victori, ныне признанные синонимичными. Точно не ясно, синонимы ли Archeria и Cricotus, но последнее имя уже не используется из-за недиагностичности материала. Чаще всего вид называют A. crassidiscus, хотя нередко также встречается похожее имя A. crassidisca. На латыни окончание -us говорит о мужском роде, -um - о среднем, -a - о женском.
Основные особенности
Средних размеров эмболомер (длина черепа - от 20 до 30 сантиметров, длина тела - до 2 метров), возможно, единственный пермский (лишь ещё один вид эмболомеров мог существовать позже арчерии - Aversor из семейства эогиринид). Как видно на фотографии, череп арчерии уплощён и имеет очень длинное рыло, хотя нижняя челюсть довольно высокая. Хорошо заметны каналы боковой линии (см. рисунок 13). В плечевом поясе присутствуют межключица и клейтрум (см. рисунок 21) - примитивная кость, имевшаяся у многих примитивных амфибий. У современных костных рыб она располагается под горлом. Кисть эмболомерного типа (см. рисунок 20), число фаланг пятого пальца - вероятно, 4. Тазовый пояс удлинённый, прочный и цельный, задние конечности сильнее передних. Самый длинный палец стопы - пятый. Позвонков (см. рисунок 22) больше, чем у остальных эмболомеров. Удлинённое тело способствовало охоте и передвижению в воде, этому помогал и хвостовой плавник, неизвестный у родственников арчерии: он был образован из так называемых супраневнальных радиалий. Радиалии у большиства рыб - костные лучи, которые поддерживают лучи кожной лопасти (лепидотрихии). Это основа скелета всех рыбьих плавников, не только хвостового. Устройство плавника на рисунке 23:
Рисунок 23. Хвостовой
плавник рыбы
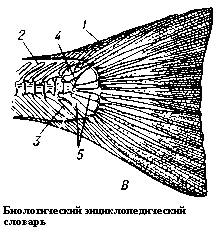
Термины: радиалии (4); лепидотрихии (1)
______________________________________________________________________________________________
Archeriidae: Cricotus: С. heteroclitus COPE 1875
Рисунок 24. Череп Cricotus heteroclitus
В конце XIX века в США было найдено много разрозненных эмболомерных позвонков. Позвонки из Техаса Эдвард Коп отнёс к синонимичным видам C. crassidiscus и C. hypantricus (Cope 1884), а из Иллинойса - к другим синонимичным видaм C. heteroclitus, C. gibsoni и C. discophorus. Первые прировняли к Archeria (Case 1915), а C. heteroclitus остался единственным видом рода Cricotus. Помимо позвонков из разных областей позвоночника, от данного вида известны короткие и широкие фаланги пальцев, а также череп (см. рисунок 24). Ввиду его удлинённости рассматривалась возможность его принадлежности к трематозавровым темноспондилам. Глазницы расположены в его задней части.
______________________________________________________________________________________________
Archeriidae: Spondylerpeton:
S. spinatum MOODIE 1912
Самый ранний из обнаруженных представителей семейства арчериид - обитал во время позднего карбона на территории США. Однако этот вид очень плохо известен - найдены лишь 9 позвонков из хвостовой области. Впрочем, отличий от арчерии и Cricotus heteroclitus достаточно, чтобы признать Spondylerpeton отдельным родом. Среди них, например, форма позвоночного столба. Высота позвонков в два раза больше ширины, и это отличает Spondylerpeton от Cricotus. Гипоцентр относительно большой: его длина - 10 мм, высота - 10,5 мм. Плевроцентр позвонка известен только один; его длина - 11,5 мм, а высота - 20 мм. Тело позвонка имеет латинское название neurocentrum, и включает гипоцентр и плевроцентр (см. рисунок 4). Общая длина образца - 60 мм.
______________________________________________________________________________________________
Anthracosauridae: Anthracosaurus: A. russelli
HUXLEY 1863
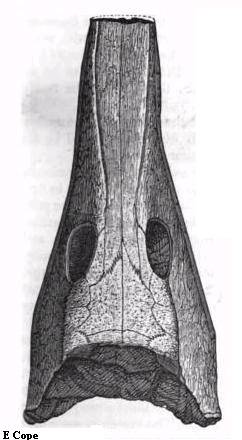
Рисунок 25. Крыша
черепа Anthracosaurus
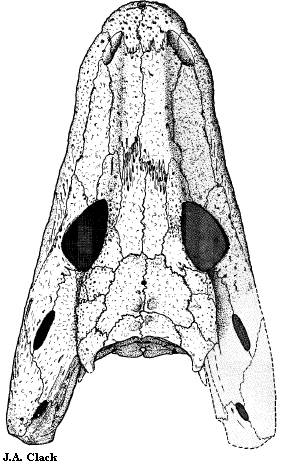
История изучения
A. russelli назван
Хаксли в 1863 году по черепу без нижней челюсти - это один из самых давно
известных эмболомеров и ископаемых земноводных вообще. Американские виды 'Anthracosaurus lancifer',
Leptophractus lancifer,
а также Rhizodus lancifer и R.
incurvus (Newberry,
1856) ныне признаны синонимичными и не относящимися к родственникам Anthracosaurus. Что
интересно, на самом деле Rhizodus
является крупной (до нескольких метров в длину) и довольно примитивной
кистепёрой рыбой, а не амфибией. Семейство антракозаврид было названо Копом (Cope 1875) и подробно
рассмотрено в работе Панчена (Panchen
1976).
Общие факты, основные
особенности
Родовое название переводится как "угольный ящер".
Видовое дано в честь маркшейдера Дж. Расселла. По имени антракозавра зачастую
называют весь отряд эмболомеров. Это ведёт к путанице, так как антракозаврами
могут называть и всех рептилиоморф (включая сеймуриаморф). Сам антракозавр -
хищное водное животное, обитавшее на территории современной Шотландии в конце
каменноугольного периода. Антракозавр являлся одним из самых крупных
представителей отряда с предполагаемой длиной тела до трёх метров, имел
массивный череп длиной почти 40 сантиметров. Вообще большинство данных об этой
амфибии, в том числе образ жизни, определёны по черепу голотипа и его отдельным
фрагментам, найденным позже; в их числе и две массивных нижних челюсти, имевших
по одному meckelian fenestra,
полученному в результате слияния двух (см. рисунок 15). Длина орбиты глаза
почти 7 сантиметров; ноздри - 3 сантиметра. Под кинетической линией, в районе
шва чешуйчатой и скуловой костей, есть окна продолговатой формы длиной 3,5
сантиметра и шириной в 1 сантиметр (см. рисунок 24). Хоаны (внутренние
отверстия ноздрей) узкие и длинные. Что касается зубов, то они хорошо
сохранились у голотипа. Длина их варьирует, самые крупные были более 3 сантиметров.
Одним из важных для систематики признаков семейства антракозаврид являются два
зуба предчелюстной кости. Посткраниальный скелет плохо сохранился. К нему
относят межключицу характерной формы, рёбра и типично эмболомерные позвонки.
Туловищный позвонок достигал около 4,5 сантиметров в диаметре. Межключица у
антракозавра больше, чем у арчерии и у другого очень крупного эмболомера - Eogyrinus.
______________________________________________________________________________________________
Anthracosauridae: Eobaphetes: E. kansensis
MOODIE 1916
Рисунок 26.
Сохранившиеся части черепа Eobaphetes
Термины: parietal (теменные кости)
История изучения
Имя Eobaphetes (эобафет, происходит от Baphetes, примитивной амфибии из группы локсоммоидов) объединяет под собой три образца. Первый был изначально назван Erpetosuchus (Moodie 1911), однако позже выяснилось, что это имя уже занято (Newton 1894). В 1916 название поменяли.
Общие факты, основные
особенности
Видовое название E. kansensis
дано по штату Канзас, где найдены окаменелости животного. В Северной Америке
найдено много образцов каменноугольных амфибий, и эобафет - одна из них. Голотип
включал некоторые районы черепа и три тела позвонков. Череп у эобафета меньше,
чем у антракозавра. На зубной кости нижней челюсти подсчитано 26 зубов. Важными
чертами семейства антракозаврид являются суммарное превосходство ширины над
длиной у теменных костей (между которыми находится теменной глаз), крупные
орбиты и массивные зубы. Различия заключаются в основном в размерах (череп эобафета
достигал 5/8 длины черепа антракозавра - почти 25 сантиметров) и в наличии у
эобафета двух meckelian fenestrae (см. рисунок 15), в то время как у антракозавра оно одно,
получившееся в результате слияния двух.
______________________________________________________________________________________________
Eogyrinidae: Leptophractus: L. obsoletus COPE 1873
и 'Anthracosaurus lancifer'
Рисунок 27. Череп Leptophractus
История изучения
Rhizodus lancifer был описан Ньюберри в 1856 году по единственному зубу. Так как и у кистепёрых рыб, и у эмболомеров имеется дентиновая складчатость зубов, они были спутаны. После этого видовое название lancifer стало применяться и к Anthracosaurus, и к Leptophractus (Romer 1930). У Панчена в работе 1976 года в семейство Anthracosauridae входит лишь Anthracosaurus russelli, а семейство Eogyrinidae разделено два на подсемейства: Eogyrininae и Leptophractinae. Последнее образуют Leptophractus, 'Anthracosaurus lancifer', Neopteroplax, а также Eobaphetes, Spondylerpeton и Cricotus heteroclitus. Такая необычная классификация строится на основе следующих признаков Leptophractinae: округлые орбиты; массивные выступы таблитчатых костей (см. рисунок 1); зубы короткие, конические и с выемками; "клыки" исключительно массивны; крупные зубы на предчелюстной кости.
Основные особенности
Все вышеперечисленные признаки применимы не только к Leptophractus (лептофракт), но и к другим членам его подсемейства. Что же касается собственно лептофракта, то от него известно несколько образцов, включая череп без задней части (см. рисунок 27). Отличительные признаки вида: относительно высокий череп длиной немногим более 10 сантиметров (следовательно, полная длина тела - около 1 метра); маленькие глазницы; хоаны крупные, в то время как наружные ноздри малекькие; глубокая ушная вырезка (см. рисунок 6); зубы очень крепкие. Самый крупный зуб лептофракта достигал длины 2,3 сантиметра.
______________________________________________________________________________________________
Eogyrinidae: Neopteroplax: N. conemaughensis
ROMER 1963 и N.relictus
Рисунок 28.
Обнаруженные части черепа Neopteroplax
conemaughensis
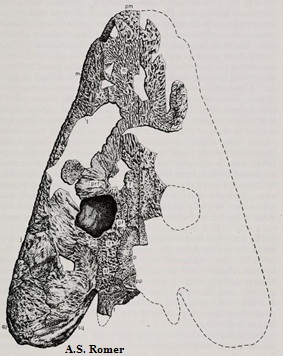
Рисунок 29.
Предполагаемый вид черепа того же вида сбоку
Общие факты
Два вида Neopteroplax (неоптероплакс) были описаны Ромером в 1963 году. Вид N. relictus назван по крупным фрагментам челюсти и позвоночника времён конца карбона и начала перми Техаса (США). Типовой вид, N. conemaughensis, из той же страны. Видовое название он получил по местности Конемауг (штат Огайо), где были обнаружены окаменелости, а родовое (новый Pteroplax) произошло от имени другого эмболомера, который будет рассматриваться позже. Время существования типового вида – поздний карбон.
Основные особенности
Основную часть материала N. conemaughensis составляет хорошо сохранившаяся левая часть крыши черепа, а из посткраниальных останков известны лишь некоторые фрагменты рёбер и позвонков (диаметры тел которых около 3 - 3,5 сантиметров). Максимальная длина черепа от рыла до крайних квадратных костей (на рисунке 29 - q) составляет почти 40 сантиметров, следовательно, полная длина тела животного около 3 - 4 метров. Таким образом, это один из крупнейших эмболомеров, уступающий в размерах только Anthracosaurus russelli и Eogyrinus, а также, возможно, Pteroplax с похожими пропорциями черепа. Как и большинство представителей отряда, неоптероплакс имел 2 больших meckelian окна в нижней челюсти, однако на его черепе не найдены каналы боковой линии. Предчелюстная кость (pm) толстая, в то время как верхнечелюстная (m) удлинённая и узкая. Последняя достигает почти 24 сантиметров, то есть более половины длины черепа. Слёзная кость (l), находящаяся рядом с верхнечелюстной, узкая в передней части. Носовая кость (n), соприкасающаяся с предчелюстной и слёзной, очень крупная. Заглазничная кость (po) напротив, небольшая. Что же касается зубов, то их крайний ряд на верхней челюсти (как и на нижней) включает около 40 штук. Из трёх зубов на предчелюстной кости первые два особенно крупные – их диаметр равнялся примерно 1 сантиметру. Максимальный диаметр крайних зубов неоптероплакса – до 1,5 сантиметров, средний – 0,6 сантиметра. В то же время, нёбные клыки были намного больше и достигали 2,5 сантиметров в диаметре.
______________________________________________________________________________________________
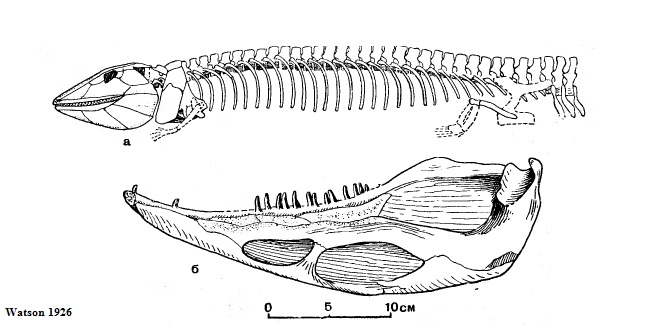
Eogyrinidae: Pteroplax: P. cornutus HANCOCK et ATTHEY 1868
Рисунок 30. Реконструированный
скелет Pteroplax без хвоста и нижняя челюсть с
внутренней стороны
История изучения
Единственный вид рода Pteroplax (птероплакс) – P. cornutus (часто встречается и написание P. cornuta) был одним из первых описанных эмболомеров – о нём узнали ещё в позапрошлом веке. Иногда название этого рода считается синонимом двух других названий родов эогиринид – Eogyrinus и Pholiderpeton, по крайней мере, строением очень похожих на птероплакса. Птероплакс фигурировал также в работах Ватсона (1926) и Ромера (1963), а более чем через столетие после оригинальной работы был описан осевой скелет птероплакса (Boyd 1980).
Общие факты, основные особенности
Довольно крупный шотландский эмболомер, живший во время позднего карбона, примерно одинаковой длины с американским неоптероплаксом. Известны несколько образцов, включая части черепов, один из которых был принят изначально за череп антракозавра, а также позвонки и рёбра. Длина черепа птероплакса – около 40 сантиметров, значит, полная длина животного – 3 – 4 метра. Как и другие эогириниды, имел большие meckelian окна в нижней челюсти. Пропорции черепа в общих чертах схожи с таковыми у неоптероплакса. У птероплакса менее округлые орбиты по сравнению с неоптероплаксом, ещё более высокая нижняя челюсть (это видно по рисункам). На предчелюстных костях по пять или шесть конических зубов, самые крупные длиной превышали 1 сантиметр. Есть и другие признаки, по которым птероплакс ближе к английским эмболомерам – Eogyrinus и Pholiderpeton, чем к североамериканским лептофракту, неоптероплаксу и прочим, образующим редко использующееся подсемейство Leptophractinae. Плевроцентры птероплакса почти вдвое толще гипоцентров. Средний диаметр предкрестцовых позвонков – чуть более 2 сантиметров. Интересная особенность невральных дуг данного эмболомера – наличие надневрального канала (см. рисунок 31 - sc). Интересно, что похожий канал имелся не только у близкого к птероплаксу Eogyrinus, но и у арчерии.
Рисунок 31. Позвонок Pteroplax
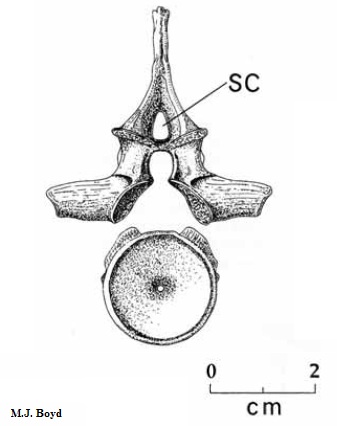
Термины: надневральный канал (supraneural canal)
______________________________________________________________________________________________
Eogyrinidae: Eogyrinus: E. attheyi WATSON 1926
Рисунок 32. Нижняя
челюсть Eogyrinus
Рисунок 33. Нёбо Eogyrinus
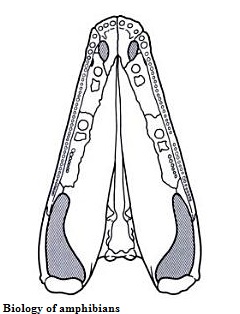
Общие факты, основные
особенности
Eogyrinus (эогирин) был назван Ватсоном в 1926 году. Родовое имя означает «ранний/рассветный головастик», а видовое дано в честь палеонтолога, открывшего птероплакса. Известен по неполным образцам: Панченом сначала описан осевой скелет (1966), а затем череп вместе с межключицей, бедром и брюшными чешуйками данной амфибии (1972). Возможно, Eogyrinus является младшим синонимом Pteroplax и Pholiderpeton (фолидерпетон), открытых ещё в XIX веке. Все три рода из позднего карбона севера Великобритании: птероплакс из Нортамберланда, эогирин оттуда же, а также из Шотландии, и фолидерпетон из Йоркшира (самый южный из трёх видов). Реконструкция скелета эогирина мало отличается от таковой у птероплакса (см. рисунок 30), поэтому нет смысла её здесь приводить. Эогирин считается самым большим эогиринидом из известных, а, возможно, и крупнейшим эмболомером; впрочем, если он и превосходит других длиной, то не намного. Полная длина черепа эогирина (от рыла до квадратных костей) - 43 сантиметра, а средняя линия черепа типового образца (длина от рыла до затылка) - 34 сантиметра: у птероплакса и неоптероплакса данные показатели лишь немного меньше. Диаметр теменного глаза всего 2 миллиметра. Позвонки похожи на позвонки птероплакса: тонкий гипоцентр, толстый плевроцентр и надневральный канал. Имел одно характеризующее крестцовое ребро, а первые несколько хвостовых позвонков несли рёбра необычной формы. Большое количество позвонков, а, значит, и вытянутость тела, определяло образ жизни эогирина и его ближайших родственников – крупные водные охотники.
______________________________________________________________________________________________
Eogyrinidae: Pholiderpeton: P. scutigerum HUXLEY 1863 и P. bretonense ROMER 1958
Рисунок 34. Голотип P. scutigerum
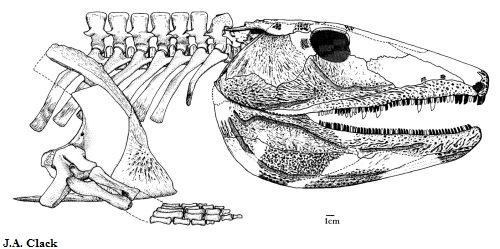
Рисунок 35. Весь скелет (предполагаемый)
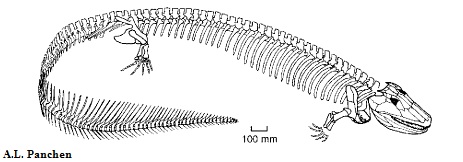
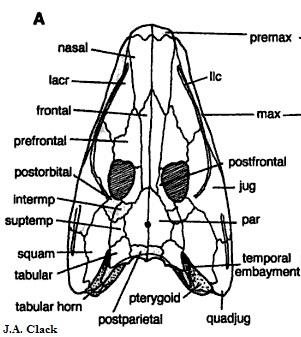
Рисунок 36. Череп (вид сверху)
Общие факты, основные
особенности
Pholiderpeton scutigerum ("чешуйчатый гад") - один из первых найденных и описанных эмболомеров. Известен по передней части скелета, включая череп и передние конечности. Строением сильно похож на эогирина, поэтому многие авторы, в первую очередь Дж. Клак, объединяют их в один род, причем Pholiderpeton является старшим синонимом. Схожих признаков очень много, поэтому здесь перечислим наиболее заметные различия:
|
Pholiderpeton scutigerum |
Eogyrinus attheyi |
|
Максилла заканчивается до задней границы орбиты |
Максилла продолжается за задней границей орбиты |
|
Задняя грань скуловой не заострена |
Задняя грань скуловой заострена |
|
Скуловая заканчивается до передней границы орбиты |
Скуловая продлевается спереди передней границы орбиты |
|
Пинеальное отверстие за серединой шва между теменными |
Пинеальное отверстие перед серединой шва между теменными |
|
Сошник с парой клыков |
Сошник без пары клыков |
|
Зубная с парой клыков спереди |
Зубная без пары клыков спереди |
|
Преартикулярная не сшита с крышечной |
Преартикулярная сшита с крышечной (нижняя челюсть) |
|
Задний короноид выставлен на виде сбоку |
Задний короноид не выставлен на виде сбоку |
Все признаки взяты из работы (Ruta, Coates and Quicke 2002), но возможно выделить ещё один признак. Сравним рисунок 33 и следующий:
Рисунок 37. Череп Pholiderpeton
По рисунку 37 видно, что птеригоид фолидерпетона (подписан на виде снизу) не соприкасается со скуловой костью, в то время как у эогирина (рисунок 33) аналогичные кости соединены швом. Это можно считать ещё одним различительным признаком, но он не внесен в таблицу, так как в вышеупомянутой работе данные другие. Рисунки взяты из достоверных источников, и тем более странно это противоречие.
Ещё одна заметная особенность фолидерпетона – наличие аноклейтрума (как у тулерпетона) – примитивный признак; имелся ли он у эогирина – неизвестно. Но в общем итоге, различий между этими двумя видами довольно много. М. Рута вообще разбивает семейство эогиринид и объединяет фолидерпетона с арчерией, а эогирина с антракозавром (их роднит, в частности, предпоследний признак из таблицы). Помимо тонкостей строения черепа, перечисленных выше, фолидерпетона от эогирина отличает также размер: по разным оценкам, фолидерпетон был от 1 до 3 метров в длину, а эогирин – не менее 4 метров. Найдены оба в Великобритании, хотя и в разных местах (эогирин – в Шотландии, фолидерпетон – в Йоркшире), зато время обитания обоих абсолютно одинаковое (башкирский ярус верхнего карбона).
Рисунок 38. Ключица P. bretonense
Что касается Pholiderpeton bretonense, то это сомнительный вид, о котором очень мало известно. Описан по большей части нижней челюсти из Новой Шотландии (Канада); возможно этот вид несколько старше, чем остальные эогириниды. Кроме того были найдены другая нижняя челюсть, чуть меньше голотипа, но похожая (возможно, принадлежала молодой особи), а также ключица (см. рисунок), предположительно относящаяся к этому виду. Учитывая скудность информации и местонахождение окаменелостей, есть основания полагать, что P. bretonense на самом деле может не относиться к роду фолидерпетон.
РАБОТА НАД СТАТЬЕЙ ОСТАНОВЛЕНА НА НЕОПРЕДЕЛЕННЫЙ СРОК
ССЫЛКИ НА ВСЕ ИСТОЧНИКИ ИЛЛЮСТРАЦИЙ УКАЗАНЫ.